|
<解説記事ダウンロード>
<概要>
アポトーシスは、生理的、病理的要因により生じた不要な細胞や障害細胞などを積極的に除去する能動的細胞死である。形態形成や恒常性維持、生体防御機構として、細胞増殖や分化などと表裏一体となって働き、多細胞社会たる生体の維持に必須の役割を演じている。したがって、その異常は、癌、自己免疫疾患、発生異常他の多岐に亘る疾病と関係する。研究の進展に伴い、アポトーシス発現の分子機構の多様性が次々に明らかになっている。放射線誘発アポトーシスは、細胞の種類や状況により大差があり、照射後分裂を介さずに起こる間期死型と、数回の分裂を経て生じる増殖死型があり、その発現機構も多様である。この細胞死は放射線障害の発現と抑制の両面に働き、また、放射線癌治療にも関与する。 <更新年月> 2006年10月
<本文>
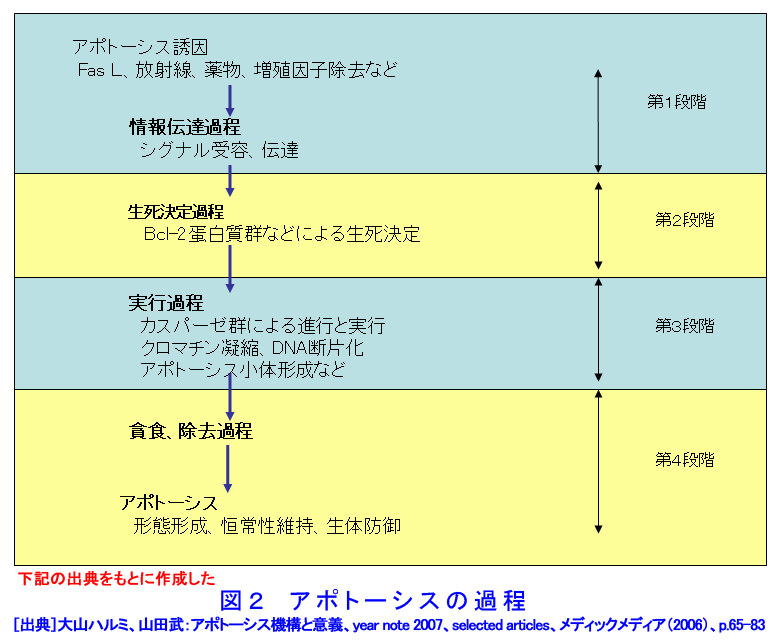
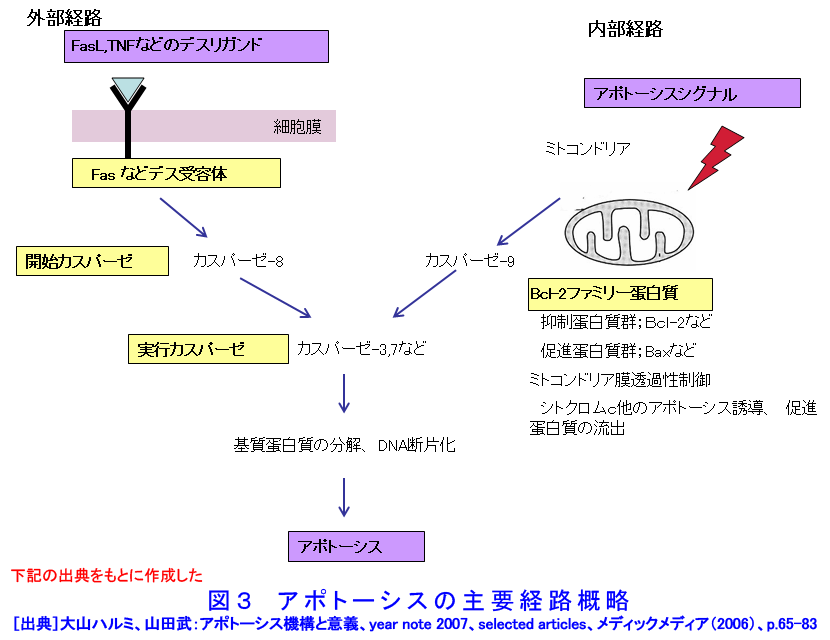
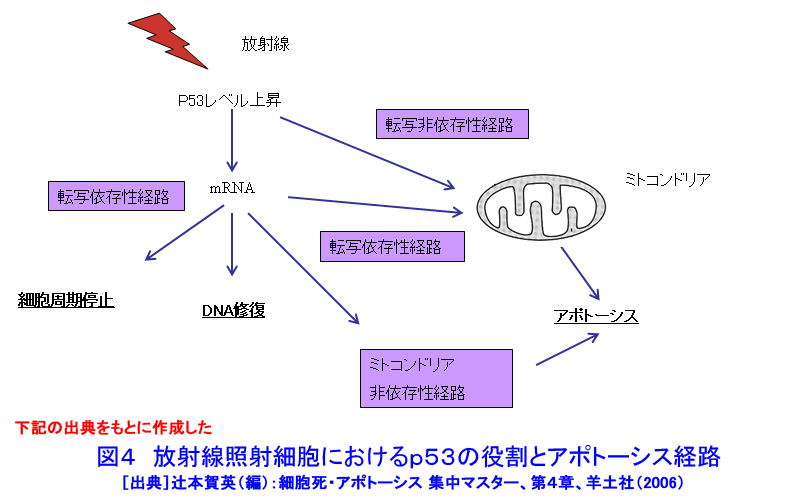
1.アポトーシスとは(参考文献1〜3) 長い間、細胞死とは病理的細胞死であるネクローシスを指していた。1972年、Kerrらはネクローシスとは形態学的に異なり、細胞縮小やクロマチン凝縮などの特徴的な変化を示す細胞死が多くの生理的、病理的な要因により生じることを見いだし、その生体制御機構としての重要性を強調しアポトーシス(apoptosis)と名付け、画期的な細胞死の概念を提起した。 ネクローシスは虚血などの病理的な要因により一群の細胞に一様に生じ、徐々に細胞膨化などが進行し、炎症を伴う。一方、アポトーシスは様々な誘因により、一旦開始すると急速に細胞縮小、blebbingという大小の球状突起の出現、クロマチン凝縮などが起り、次いで球状突起が断片化してアポトーシス小体となり、速やかに貪食除去される。この細胞死は散発的に起こる特徴があり、炎症は生じない(図1)。 アポトーシスの特徴的な生化学的変化の一つがクロマチンDNAのオリゴヌクレオソーム単位の切断によるDNA断片化(DNA fragmentation)である。アポトーシスの重要な指標の一つとして使われている。アポトーシスは進化の早期に獲得された細胞の基本的な機能であり、わずか約1000個の細胞しかない1ミリの線虫における細胞死が、アポトーシスと相同な機構で起こっており、実際、アポトーシス分子機構の研究の進歩は、線虫の細胞死機構の研究に負うところが大きい(参考文献2〜5)。 2.アポトーシスの発現機構(参考文献2〜9) アポトーシスは細胞の種類や誘因によらず形態学的にほぼ同様な変化を示すが、その発現の過程は、1)アポトーシスの情報伝達、2)生死決定過程、3)実行過程、4)貪食除去過程に大別される(図2)。 アポトーシスの第1段階を開始させる誘因は、デスリガンドというFasリガンドやTNF、カルシウム、ホルモン、増殖因子などの欠如、放射線、薬物、ウイルスなど多様である。それらの情報はFas(CD95/AP0-1)、TNFRなどの受容体で受信、あるいはDNA損傷を検知、セカンドメッセンジャーで伝えるなど対応する多数の経路で伝達される。情報伝達路は、死の信号ばかりでなく、細胞の増殖、分化の情報回路とも交叉する部分が多く、この細胞死は機能、分子機構の両面で、細胞分裂とともに多細胞社会たる生体のバランスを維持する調整役となっている。また、アポトーシスは、RNAや蛋白質合成依存性に起こる場合が多く、能動的死であるとの意から、“細胞自殺”とも言う。第2段階は、生死の決定過程であり、多数のBcl-2ファミリー蛋白質などが生死決定に働く。3段階は特異的なプロテアーゼであるカスパーゼが多様な蛋白質を限定分解し、アポトーシス過程の進行と死に伴う特徴的諸変化を起こし細胞自壊に導く。第4段階では、アポトーシス小体が近隣細胞やマクロファージにより貪食、除去される。 これらの過程には複数の経路があり、第1−3段階では大きく分けて主な2経路は、1)FasL/Fasなど死のシグナルと受容体を介する外部経路と、2)ミトコンドリアのBcl-2ファミリー蛋白質により制御される内部経路で、両経路共にカスパーゼが多段階に働き進行する(図3)。外部経路は主に免疫系の機能保持に働き、内部経路は主に放射線やストレスなど多くの要因に誘導される。 Bcl-2ファミリー蛋白質は、現在30以上の分子が発見されており、アポトーシスを抑制する蛋白質群と促進蛋白群がある。これらは主にミトコンドリアの膜透過性を調節し、シトクロムcやアポトーシス進行に働く蛋白質のミトコンドリアからの流出を制御、抑制蛋白質と促進蛋白質のバランスが細胞の生死決定に働く。 カスパーゼはアスパラギン酸に隣接する末端部位Asp-X間で蛋白質を切断、限定分解する特異的なプロテアーゼで、現在まで14種発見されており、各カスパーゼにより切断する基質が異なる。通常は不活性型の前駆蛋白質として存在し、自己あるいは他のカスパーゼにより分解され活性型となる。カスパーゼはシグナルを受けてから上流で働くイニシエーター(開始)カスパーゼ(外部経路では主にカスパーゼ8、内部経路ではカスパー−9)と、それらにより分解され活性化して死の執行に働くエフェクター(実行)カスパーゼ(カスパーゼ3、7など)に分けられる。 カスパーゼによる分解の基質となる蛋白質は300以上報告されており、アクチン、ラミンなどの分解による形態変化、ポリ(ADP−リボース)ポリメラーゼ(PARP)などDNA修復分子の失活、Bcl-2、Bcl-XLなどアポトーシス抑制蛋白質の失活等々を引き起こす。DNA断片化に働く酵素の一部の活性化にも、カスパーゼが関与する。カスパーゼはアポトーシスを進行させ、さらに、DNAを分解し、修復・再結合能を無くし遺伝子としての働きを喪失させ、形態変化を起こし、また、多くの酵素を失活させるなど、死細胞を無害で貪食されやすい形にまで処理する死の実行の主役である。 p53は約半数の癌で欠失や変異が認められている癌抑制遺伝子であり、p53蛋白質は放射線を含むDNA損傷などストレスに対する応答に多彩な役割を演じている。これらストレス要因によりp53レベルが上昇し、転写因子としてきわめて多数の遺伝子の発現制御に関わり、細胞周期停止、DNA修復あるいはアポトーシスを引き起こす。p53変異や欠失により、生物個体、癌を含む細胞の放射線抵抗性が増加することが多い。p53によるアポトーシス誘導には、多くの転写依存性の標的遺伝子産物を介する複数の経路に加え、最近、直接ミトコンドリアに働く転写非依存性経路も報告されている。これらは、主にミトコンドリアを介する経路で進行する。なお、放射線はDNA損傷要因としてのみでなく、膜や細胞質にも働いてアポトーシスを誘発する経路もある(図4)。p53非依存性の経路も複数存在し、また、p53依存性および非依存性のFasが関与する経路もある(参考文献3〜9)。 3.アポトーシスの意義と放射線影響(参考文献1〜11) アポトーシスは、発生過程のきわめて初期から検出され、多様な器官の形成過程において、不要となった細胞や異常細胞を除去し、形態形成に必須の働きをしている。異常胚を流産させて奇形発生を抑制し、その異常は、奇形や発育不全、あるいは、未分化細胞の残存による癌や良性腫瘍の素因ともなる。 成体においても、造血系や消化管上皮、皮膚、精巣など細胞再生系で活発な細胞増殖が行われ、こうした臓器では一般に必要数より多くの細胞が作られ、成長因子の制御などにより正常な組織構築に要する以外の過剰な細胞や、役目を終えた細胞はアポトーシスで除去される。増殖と細胞死のバランスの不均衡が、臓器萎縮や過形成を引き起こし、50%以上の疾病にアポトーシスが関与しているとされる。アポトーシス機構の回避、過少は癌、自己免疫性疾患、ウイルス感染など、逆に過剰なアポトーシスは、AIDS、アルツハイマー病、血液疾患、放射線障害、神経変性疾患など多くの疾病と関連する。 放射線照射による正常細胞のアポトーシスは、多様な障害の原因となる。放射線事故の場合や、治療の際の副作用の点から各種の正常細胞の放射線誘発アポトーシスが問題となる。逆に、癌治療の場合には、アポトーシス誘導能との関連が注目される。 放射線照射は障害や症候が検出されない線量でも、多くの細胞にアポトーシスを起こしている。たとえば、小腸クリプトの特定の幹細胞はわずか数cGyのX線やγ線照射後短時間でアポトーシスを起こし、1Gyで完全に消失する。しかし、より抵抗性の幹細胞が増殖、代替するため、10Gyまでは致死的障害が出ない。こうしたアポトーシスは、DNA損傷細胞を丸ごと除去し、酵素的に行われるDNA修復のミスを回避する高次の修復機構として働くと考えられている。DNA損傷細胞を積極的に自殺除去させる“流産型組織修復(Abortive repair)”あるいは、“利他的細胞死(Altruistic cell death)”とも呼ばれ、異常DNAを持つ細胞の増殖による遺伝的障害や発癌などのリスク抑制に寄与する。過度のアポトーシスは放射線障害の原因となる。 形態形成前期までの照射では高率に流産が起こり、奇形はほとんど生じない。ところが、形態形成期の照射では、高率な奇形が発生する。例えば、指肢形成期時には、正常でも指の間の細胞がアポトーシスを起こしヒトやマウスでは水掻きがない。しかし、その時期の照射では、指となる原基細胞にもアポトーシスを誘発し、高率に奇形を起こす。この過程にも、p53が関与し、ノックアウトマウスではアポトーシスと奇形の低下が認められる(図5)(参考文献10)。 放射線による細胞死研究は、放射線生物学研究の初期から行われ、照射後、分裂を介さずに起こる“間期死”と、コロニー形成能の低下で判定される“増殖死”に分けられてきた(放射線の細胞に対する影響の項参照)。間期死を起こす細胞は比較的少なく、胸腺細胞や小腸クリプト細胞の一部などがあり、一方、多くの細胞は数回の細胞分裂を経てから長時間後生じることが観察されている。増殖死は細胞の死そのものを見ているのではないが、癌治療の指標として有用であり、コロニー形成能の測定は今でも重要な放射線効果の判定法として使われている。 アポトーシス研究の多くの基礎的知見は、胸腺細胞を用いた研究から得られた。胸腺細胞は数Gyの照射、数時間でアポトーシスを起こす。癌細胞の中にも間期死型のアポトーシスを起こす細胞もあるが、増殖死型のアポトーシスを起こす細胞も多い(図6)(参考文献11)。アポトーシスを誘発する放射線の線量は、細胞による差がきわめて大きく、100Gyでも起こさない細胞もあり、また、同一細胞でも細胞周期など状況によって変化する。アポトーシス誘発時間も1−2時間から100時間以上と大差がある。長時間後のアポトーシスは、この死が散発し、速やかに貪食除去される特徴のため検出しにくい。放射線誘発アポトーシスの発現機構は多様である。アポトーシス誘発に対する感受性差の決定要因や、多様な機構の詳細については未だ不明な部分が多い。また、増殖死とアポトーシスの関連についてもよくわかっていない(参考文献9〜11)。 アポトーシスは、ホルミシスなども含めた多岐にわたる放射線影響に、広範に関与しているとされる。今後、アポトーシス感受性の違いの解明や放射線影響との関連研究の進展、さらには、アポトーシスの促進と抑制に働く薬剤や方策の開発が進めば、放射線障害抑制と癌治療などに資することが期待される。 <図/表> 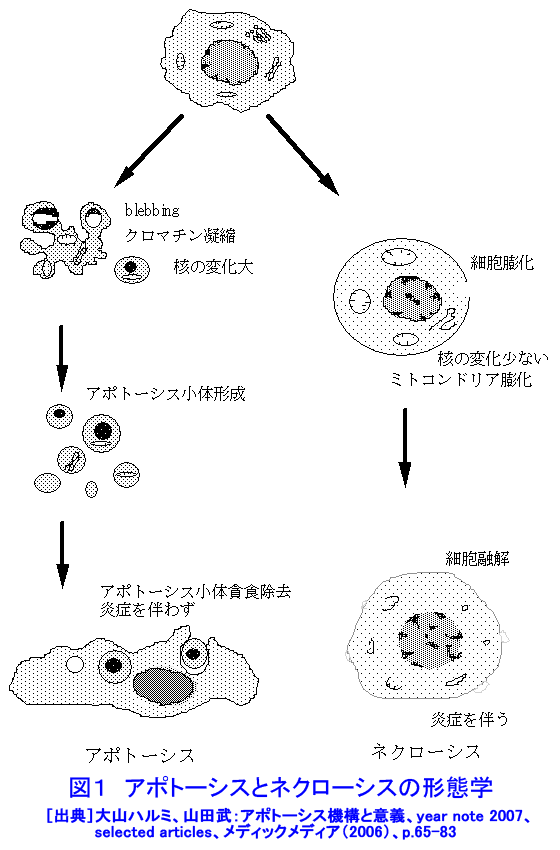
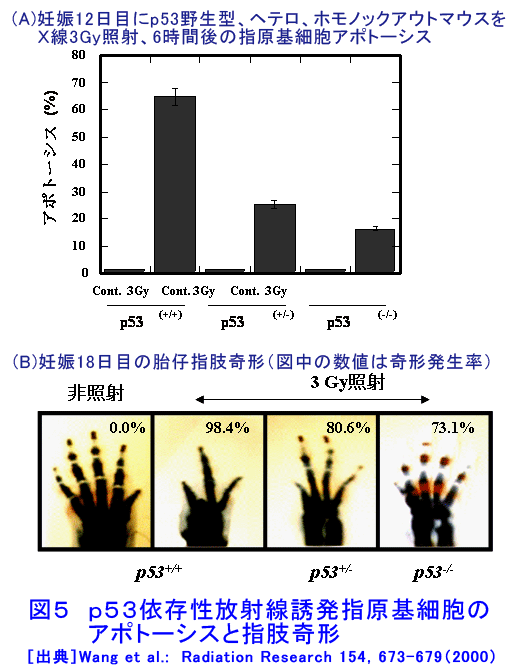
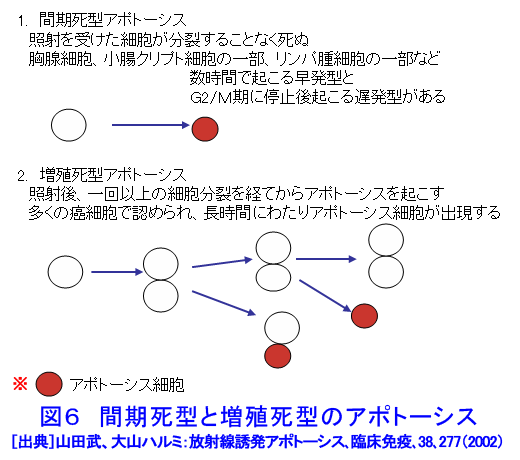
<関連タイトル> イオンビームバイオ技術の現状 (08-03-01-07) 放射線と生物の適応応答 (09-02-01-02) 放射線ホルミシス (09-02-01-03) 放射線の細胞への影響 (09-02-02-07) 放射線効果と修復作用 (09-02-02-12) 放射線の細胞分裂に及ぼす影響 (09-02-02-16) 放射線の晩発性影響 (09-02-03-02) <参考文献> (1)Kerr,J.F.R.,Wyllie,A.H. and Currie,A.R.:Apoptosis:a basic biological phenomenon with wide ranging implication in tissue kinetics. Brit. J. Cancer 26 239(1972) (2)山田武、大山ハルミ:アポトーシスの科学、講談社ブルーバックス(1994) (3)田沼靖一(編):アポトーシスがわかる、わかる実験医学シリーズ、羊土社(2001) (4)大山ハルミ、山田武:アポトーシス機構と意義、year note 2007、selected articles、メディックメディア(2006)、p.65-83 (5)辻本賀英(編):細胞死・アポトーシス 集中マスター、羊土社(2006) (6)月刊「臨床免疫」編集委員会(編):アポトーシスのすべて、臨床免疫、第38巻特別増刊号、科学評論社(2002) (7)辻本賀英、一條秀憲(編):成熟・展開するアポトーシス研究、実験医学増刊号、Vol.22、No.11(2004) (8)條秀憲(編):特集 アポトーシス 分子メカニズムとその破綻が引き起こす疾患 、Molecular Medicine、Vol.39、No.6(2002) (9)Potten C. and Wilson J.:Apoptosis The Life and Death of Cells. Cambridge University Press.(2004) (10)Wang et al.: Radiation Research 154,673(2000) (11)山田武、大山ハルミ:放射線誘発アポトーシス、臨床免疫、38、277(2002)
|

