|
<解説記事ダウンロード>
<概要>
中性子イメージングプレート(中性子IP)は、輝尽性蛍光体(PSL: photostimulated luminescence、Euをドープしたバリウムフルオロハライド(BaF(Br,F))に中性子コンバーター(Gdないしは6Li)を均一に混入させた積分型エリア熱中性子検出器である。ダイナミックレンジは5桁、位置の分解能は200μm以下、中性子検出効率は50〜100%(中性子波長に依存)の性能を有し、1mm以下厚の柔軟で取扱い容易なフィルム状(大きさ20cm×20cm以上)のものである。 中性子ラジオグラフィ、タンパク質等の生体物質の水素原子位置を含めた三次元立体構造解析、その他粉末中性子回折、中性子小角散乱、中性子反射率計等多岐にわたる測定、解析に応用されている。 <更新年月> 2006年08月
<本文>
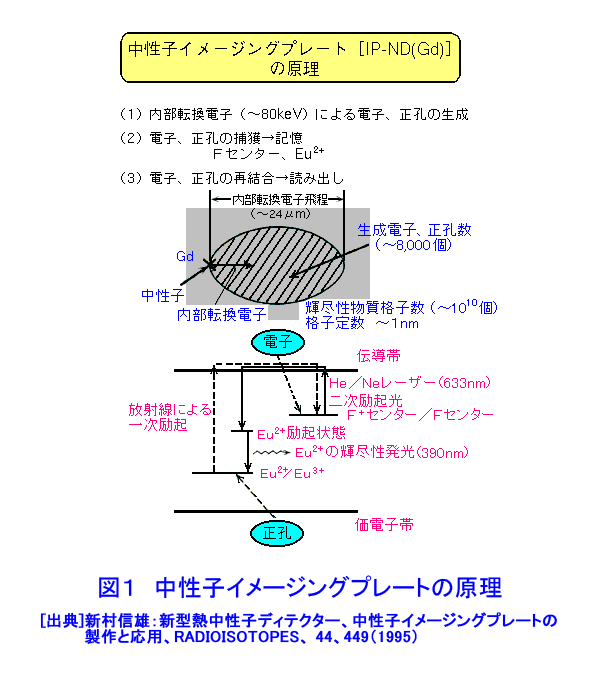
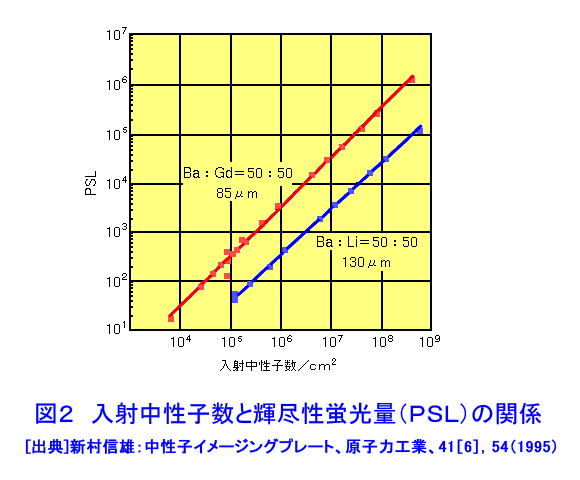
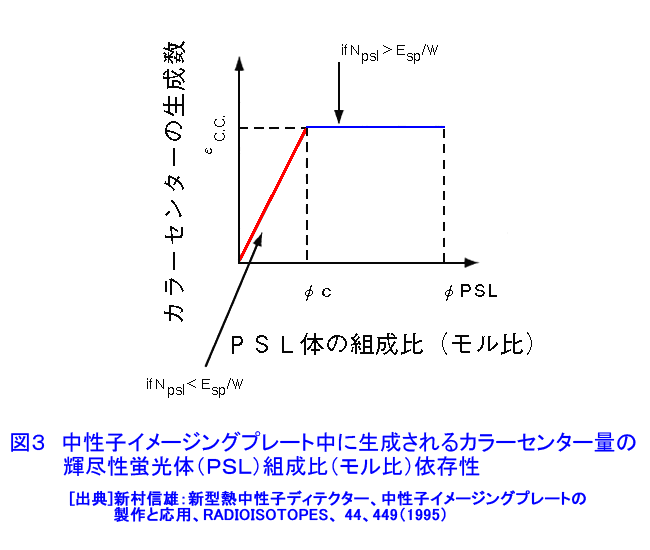
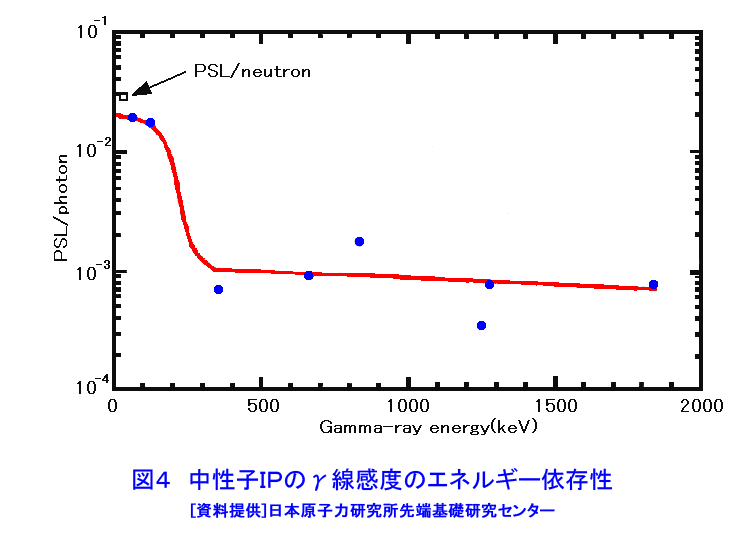
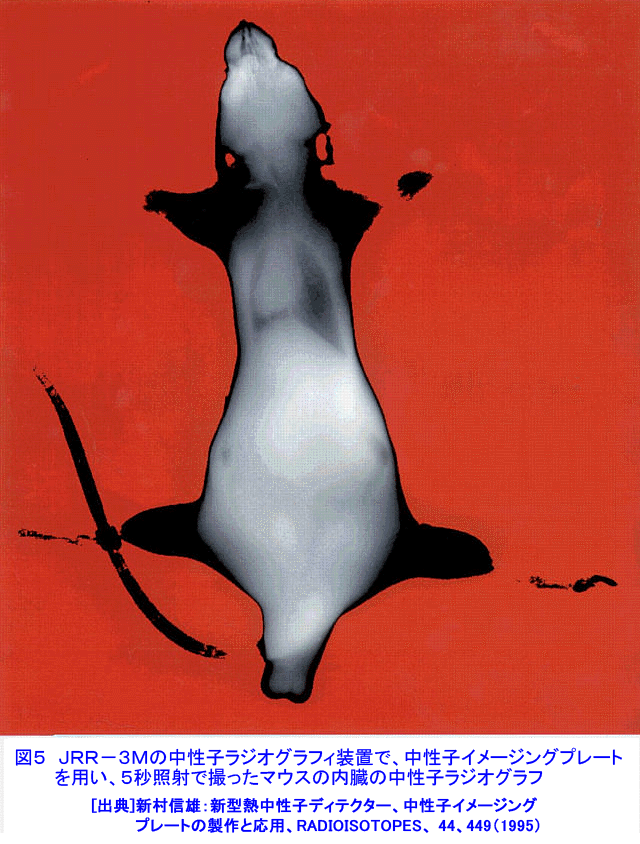
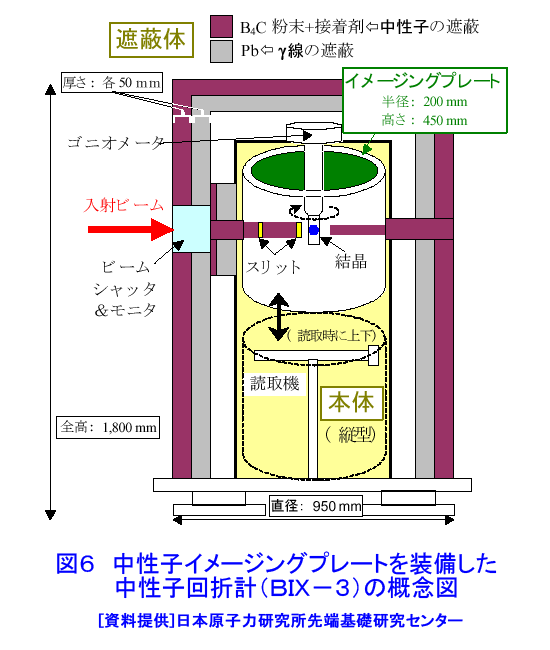
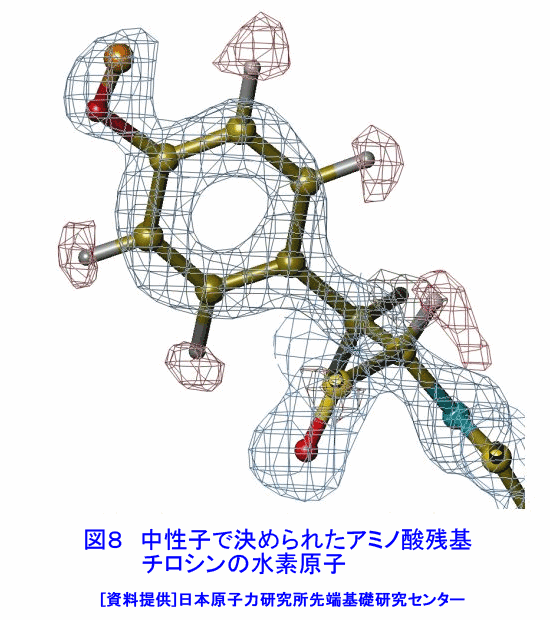
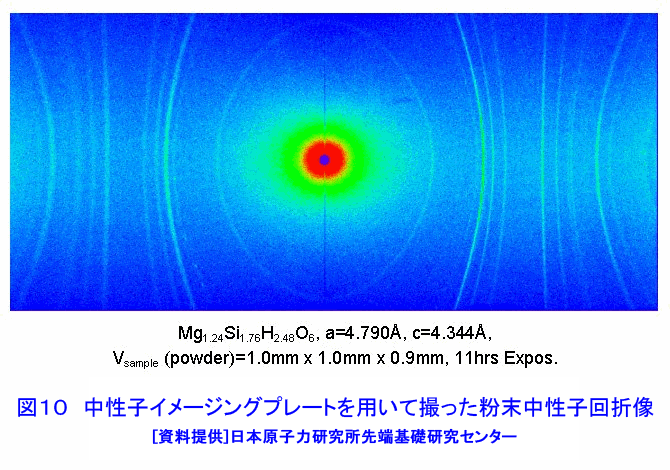
中性子は、電離作用はないが、熱中性子は、ある核種と核反応をするので、そのとき生じる二次荷電粒子の電離作用を利用して、中性子を検出することができる(これらの核種を中性子コンバーターと呼ぶ)。たとえば10B、3Heは気体比例計数管、6Liはシンチレータ、Gdは写真法、そして235Uは核分裂検出器に用いられている。最近は、中性子入射位置を弁別できる気体比例計数管エリアディテクターが多く使われているが、その標準的な性能は、検出効率:50〜90%(中性子波長に依存する)、最大計数率:1.0E4〜1.0E6カウント/秒、位置分解能:1〜5mmが原理的な限界であった。そのため、1桁以上の位置分解能等の性能向上には新しいタイプの検出器の開発を待たざるを得なかった。 このような背景の下で、中性子イメージングプレート(IP)が開発された(参考文献1)が、熱中性子検出器としての性能を総合的に検討すると、中性子IPは中性子検出器として、未だかつて経験したことのない優れたものであることが判明し、いくつかの応用例が報告され実用に供されている(参考文献2,3,4,5,6,11)。今や中性子IPは中性子科学を支える基盤技術の一つ(参考文献15)として認識され、世界的に著名な放射線の教科書にも紹介されるようになった(参考文献12)。 1.中性子IP 1.1 原理と構造 中性子IPは、輝尽性蛍光(PSL:photostimulated luminescence)体に中性子コンバーターを均一に混入させた積分型エリア中性子検出器である。中性子コンバーターとしては、Gdないしは6Liが用いられている(図1)。中性子がコンバーターに捕獲されると、二次荷電粒子が放出される。PSL体として、EuドーピングBaF(Br,I)が用いられているが、この二次荷電粒子がPSL体中にカラーセンターを生成する。このカラーセンターは準安定で、He-Neレーザー赤色光(波長633nm)の照射で紫色発光(波長390nm)し、初期安定状態に戻る。この紫色光を光電子増倍管で検出する。その光量が入射中性子数に比例するので中性子検出器として使える。構造は、支持層としてのPET(ポリエチレンテレフタレート;180〜300μm)のシートにPSL体を50〜300μmの範囲で塗布し、その上に保護層(3〜10μm)をおいた、全体でも1mm以下厚の柔軟で取扱い容易なフィルムである。 1.2 性能 中性子IPへの入射中性子数と、それにより発光する輝尽性蛍光量(PSL)との直線関係が保たれる範囲をダイナミックレンジと呼ぶ。図2に入射中性子数とPSLの関係を示す。広範囲にわたって良い直線性を示しており、Gdコンバーターでは5桁、Liコンバーターでは4桁のダイナミックレンジを確認できた。中性子IPの位置分解能は、200μm以下で、実験条件、読取条件を適切に選べば50μm程度まで可能である。中性子IPへの入射中性子はPSLとして観測にかかるが、中性子捕獲からPSL発光までにはいくつかのプロセスがあるため、観測されたPSL値から中性子数の絶対値を求めるのは困難である。そのため中性子検出効率については、計算によって求めるのが妥当である。用いる中性子波長に依るが、50〜80%が得られる。 1.3 最適化 中性子IPの成分のうちコンバーター原子数を増せば中性子捕獲は増すが、PSL体の成分比は少なくなっているので、生成するカラーセンター数は減少し、PSL光量は減る。つまり、コンバーター原子数とPSL体のモル比に最適値がある。また、IPの厚みを増すとPSLがIP自体で吸収されるので、やはり厚さにも最適値がある。それら最適値は、次のように決定される。コンバーターからの二次荷電粒子は、それのエネルギー総和をEsp、一個のカラーセンターをPSL体に生成するのに必要なエネルギーをωoとすると、(Esp/ωo)個のカラーセンターを生成する能力を有する。但し、二次荷電粒子及びそれからの電離電子の飛程でPSL体中に形成される体積中にあるPSL体の個数をNpslとすると、(Esp/ωo)とNpslの大小関係で、カラーセンターの生成表式が異なる(図3)。つまり、カラーセンターの生成数は、PSL体のモル比の増加とともに、直線的に増加し、ある臨界値以上では一定値Esp/ωoとなる。一方、中性子捕獲の割合は、PSL体のモル比φpslについて、減少関数(コンバーター原子数については増加関数)であるので、中性子入射数と発光PSL量は、φpslのある一定値に最大値がある。Gdコンバーター、6Liコンバーターにつきφpsl及び厚みを変化させ、中性子入射数及びそれによる発光PSL量を実測し、これまでの議論を基に最適値を求めてみると、夫々の最適輝尽性蛍光体組成比は、それぞれ60%、15%、厚みは両方共200μmとなった。(参考文献13) 1.4 γ線に対する感受率と遮蔽 現在の中性子IPはX線用IPにコンヴァータ(最終的に実用化された中性子IPではGdを使用した。)をドープしたものでガンマ線有感である。微分型検出器であればイヴェント毎に放射線特有の信号の特徴を利用して(例えば波高分析する等)、ある程度のガンマ線弁別が可能であるが、中性子IPは積分型検出器であるので、中性子以外の放射線はいかに僅かであっても有感であれば積分されて行きバックグランドになる。これは積分型検出器が抱えるアキレス腱である。X線用IPの輝尽性蛍光物質はX線感受率を上げるためなるべく重原子で構成されるBaFBrを用いており、それをベースに開発された中性子IPは生来的にガンマ線に有感であり、中性子IPの最大の問題点と考えられ、中性子IP のガンマ線感受率を下げる改良が精力的になされた。軽原子で構成される物質として最も試されたのがKClである。(参考文献14)それにより確かにガンマ線感受率は減少したが、報告によればKClの発光量が少なく、結局中性子検出効率、位置の分解能等で従来の中性子IP を凌ぐには至っていない。中性子IPは、γ線に有感であるので、一般にはγ線の強い放射線場での使用は不適であるが、中性子IPのγ線感度のエネルギー依存性を詳細に測定すると、十分遮蔽できることが判明した。図4に中性子IPのγ線感度のエネルギー依存性を示す。300keV以下のγ線エネルギーに対する中性子IPのγ線感度は中性子感度の半分であり、300keV以上のγ線エネルギーに対する中性子IPのγ線感度は中性子感度の1/20以下になる。中性子IPの前に1mm厚の鉛を置くと、300keV以下のγ線は殆ど吸収されるが、熱中性子は殆ど吸収されないのでこれで十分なγ線遮蔽となる。また、300keV以上のγ線の感度は著しく減少するので問題ないが、例えば、JRR-3Mクラスの原子炉実験室では、4〜5cm厚の鉛ブロックでγ線バックグランドを1/10に減少させられることも実測で判明した(参考文献7)。 中性子構造生物学用の中性子IPを用いた回折計をJRR-3M原子炉実験室内に設置したが、ここで述べた方式のγ線遮蔽を施し、γ線の強い放射線場での中性子IP使用に成功した。 1.5 ガス封入型比例計数管エリアディテクタとの比較 中性子IPの特徴をガス封入型比例計数管エリアディテクタとの比較でまとめてみる(表1)。 2.中性子IPによる応用 2.1 ラジオグラフィ 中性子ラジオグラフィの最近の傾向として、テレビ法で動画を撮像する研究がされているが、非破壊検査等のように内部構造を観察する需要は今なお少なからずあり、それにはフィルム法が有効である。中性子IPの感度は、X線フィルム法に較べ、約100倍良い。実際、これは252Cf線源からの中性子を用いて、同一試料でラジオグラフィをX線フィルム法および中性子IPでそれぞれ撮像した実験で確かめられている。ただし、ダイナミックレンジは中性子IPでは4〜5桁であるのに対し、X線フィルムでは2桁しかない。ただし、肉眼のダイナミックレンジも2桁あるので、X線フィルム上に撮った像とイメージングプレートの像を肉眼で評価するのは適当ではない。JRR-3Mに設置されている中性子ラジオグラフィ装置で、中性子IPを用い、5秒照射でマウスの内臓の中性子ラジオグラフィを撮ったものが図5である。肺、心臓、肝臓、腎臓等が同定される。同じ実験を、感度1/100のX線フィルムを用いて撮ろうとすると、500秒の照射の必要性が予測されるが、生物体に相当の放射線被ばくが生じることになり、現実的でない。 2.2 中性子構造生物学(参考文献8,11) タンパク質やDNAは、三次元立体構造を形成して初めて、生理機能を発現する。そのため、生体物質の生理機能の解明に、原子レベルでの三次元立体構造解析は必須であり、これはX線結晶構造解析により行われてきた。また、生体物質は、構成元素の約半分が水素原子であり、周囲を水に取り巻かれ、水の水素原子が生理機能に大きく寄与している。ところが、X線回折法では水素原子の位置決定は困難である。一方、中性子回折法では水素原子の位置を決定することができる。このための回折装置に中性子IPは、非常に有用である。中性子IPを装備した中性子構造生物学用の中性子回折計がフランスと日本で稼働している。一つはラウエ・ランジュバン研究所(フランス)のラウエカメラで,現在LADI-II, VIVALDIという2台が稼働している。日本原子力研究開発機構JRR-3Mには2台(BIX-3, BIX-4)が稼働している。特に、BIX-3, BIX-4は取得できるデータのS/N比、および分解能(1.5オングストローム)は世界最高である。2008年の稼働を目指してオーストラリアで新しい原子炉が建設中であるが、そこでの単結晶回折装置には中性子IPが使われることになっている。このように単結晶を用いた中性子回折装置の検出器は次々に中性子IP利用になってきている。図6にBIX-3の模式図、図7にBIX-3を用いて撮った生データの1例、およびこのデータを解析して得られたタンパク質の水素原子(図8)、水和構造(図9)の例を示す。従来の中性子検出器を用いて、正方晶系ニワトリ卵白リゾチームの結晶構造解析用データ収集には、約4〜6か月のマシンタイムが必要である。1年分のマシンタイムを全部割り当てても、1〜2個の試料のデータ収集が限度である。ところが、中性子IPの出現で、20〜30日のマシンタイムで解析に必要なデータが収集ができるようになり、1年に10〜20種類の試料の実験が可能ということは、構造生物学の研究者の多くの方に、生体物質中性子回折実験を行う機会が与えられるということである。 2.3 中性子高分解能粉末回折計への応用 粉末中性子回折計の分解能は、試料から検出器への入射中性子の分散と用いる検出器の位置分解能に強く依存する。一般には、検出器の位置分解能が不十分であるため、試料と検出器の間にファインなコリメータ(ソーラースリット)を挿入し、入射中性子の分散を制限し検出器の位置分解能を見掛け上向上させ、高分解能を実現させている。しかし、粉末試料からのブラグ反射は、試料位置を頂点にして円錐状に発せられるが、幾何学的に精度の高い円錐状ソーラースリットの製作は困難であるため、一般に平行平板状ソーラースリットで代用されるのが通常で、このため検出効率は著しく低下する。一方、中性子IPのように高分解能検出器を用いると、試料の大きさ及び試料・検出器間距離を適当に選ぶだけで十分な装置分解能が得られるので、ソーラースリットが必要でなくなり、例え、試料が小さくなっても十分な検出効率が確保される。また、装置全体も大変コンパクトになる。1mm立方の粉末試料で得られた回折図形の例を図10に示す。 2.4 応用の拡大 中性子IPでは中性子照射後に読み取りという操作があるので、回折・散乱装置に組み込んでオンラインで使用するためには、従来の装置の検出器周りを大改造する必要があり結局装置全体を更新することになる。このため、現在中性子IPが回折・散乱装置に組み込まれている例は単結晶回折装置のみである。予備的な実験や提案は、ラジオグラフィー、小角散乱実験、粉末回折実験、残留応力解析実験、反射率測定実験等で行われており、その有効性が説かれているが、現実には先の理由で予算化されないでいる。穿った見方かもしれないが、これは未だ多くの研究者が本当に中性子IPの有効性を実感として持っていないのが一つの理由と考えている。この障害を取り除くために、中性子IPを簡便な中性子検出器として、次のようにどんどん使用することを提言する。 1)装置の試料位置でのダイレクトビームプロファイル測定。これの必要性は種々のケースで遭遇する。装置のアライメント時、モノクロメータを変更(種類、角度他)した時、従来中性子感受性ポラロイドが使われているが、中性子IPの方が定量的且つ簡便である。但しオフライン読み取り器が必要であるが、JRR-3M炉室、ガイドホールともにオフライン用読み取り器は装備されているのでこれに関しては問題無い。 2)ダイレクトビームの中性子絶対値測定。中性子IPは通常の散乱・回折装置のダイレクトビームを測定する位では数え落としが無いので、絶対値校正済みの中性子IPを用意しておけばダイレクトビームの絶対値が測定できる。1)と組み合わせれば併せてダイレクトビームプロファイルが求められる。但し、フェーディング効果が中性子IPにはあるので、測定時間、測定後読み取りに入るまでの時間はフェーディング効果を考慮して行う。 3)装置に装備されている検出器の守備範囲以外でのブラグ反射探索。特にクライオスタット中に単結晶試料をセットした後で赤道線以外の反射がどこにあるかが判明すれば、実験遂行が安心である場合がある。そのようなブラグ反射探索に適している。これ以外にも遮蔽の欠損の調査等種々あるだろう。このように中性子IPを簡便に使いこなす間に中性子IPの特徴が多くの人に知れ渡り、散乱・回折装置に組み込んでの中性子IPのオンライン使用の要求が起こり、使用頻度が上がれば実用化に至る中性子IP改善に繋がると考えている。 3.問題点 3.1 中性子IP構成元素の中性子による放射化 中性子IPを構成する元素のうち、Eu、I、BrやGdは熱中性子照射で放射化され、それぞれの核種固有の半減期で壊変し、イメージングプレート上でカラーセンターを形成する。これは中性子IPのバックグランドになるが、それぞれ実測した結果、中性子入射により形成されるカラーセンターの高々1.0E-7の寄与であり、これ以上の精度を要しない実験では全く問題はない。 3.2 中性子飛行時間測定法による中性子回折散乱実験に不向き 中性子IPは、積分型検出器であり、中性子入射毎にデータ処理を行う微分型検出器ではないので、中性子入射時間を測定する中性子飛行時間法への応用はできない。 <図/表> 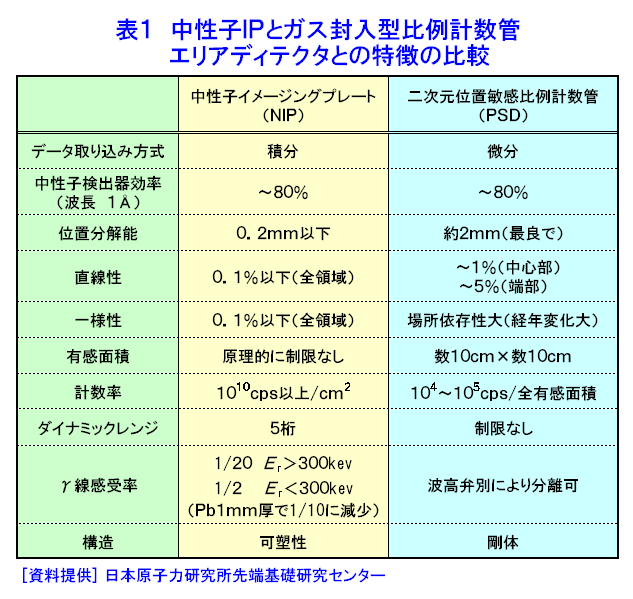


<関連タイトル> 中性子回折・散乱の原理と応用 (08-04-01-05) イメージングプレートを用いた実習実験 (08-04-01-45) <参考文献> (1) N.Niimura et al.: An imaging plate neutron detector,Nucl.Instrum & Methods A349,521(1994) (2) 新村信雄:中性子イメージングプレート、原子力工業、41[6],54(1995) (3) 新村信雄:新型熱中性子ディテクター、中性子イメージングプレートの製作と応用、RADIOISOTOPES、 44、449(1995) (4) 新村信雄:中性子検出器の革命−中性子イメージングプレートとその応用−、放射線と産業、66、41(1995) (5) 新村信雄:タンパク質中性子結晶学、日本結晶学会誌、38(1)、95-98(1996) (6) N.Niimura et. al,:Neutron Laue diffractometry with an imaging plate provides an effective data collection regime for neutron protein crystallography,Nature Struct. Biol. 4 9090914,1997 (7) Y.Haga, et. al,:Gamma-ray sensitivity and shielding of a neutron imaging plate, J. Appl. Cryst. 32,878-882(1999) (8) N.Niimura:Neutrons expand the field of structural biology, Curr. Opi. Struct. Bio.9, 602-608(1999) (9) 日本原子力研究所:蛋白質の機能発現機構の解明なるか?(2001年3月30日) (10)日本原子力研究所:研究炉利用ホームページ(2001年3月30日) (11)J.Fitter, T.Gutberlet, J.Katsaras (Eds.):Neutron Scattering in Biology. Techniques and Applications: Springer-Verlag Berlin Heidelberg 2006, p.21-72 (12) Glenn F. Knoll:Radiation Detection and Measurement (3rd ed.) 邦訳放射線計測ハンドブック第3版木村逸郎/阪井英次訳、p.842 (13) Y.K.Haga, K.Neriishi, K.Takahashi & N.Niimura:Optimization of Neutron Imaging Plate,Nucl. Instrum. Methods. 487, 504-510 (2002) (14) M.Schlapp, H.Conrad, H. von Seggern: J. Phys. D: Appl. Phys. 37, 2607-2612(2004) (15) 新村信雄:特集 中性子科学を支える基盤技術「中性子源から測定装置まで」中性子イメージングプレートの現状、日本中性子科学会誌、波紋、15、82-85(2005)
|

